基因表达的随机性,即同源细胞之间在相同基因上的表达存在差异,是生命系统中普遍存在且不可忽视的现象。在拟南芥(Arabidopsis thaliana)的花药分生组织中,这种随机基因表达尤其体现在生长素信号途径中,成为调控植物花器官形成与发育的关键调节机制。生长素是一类植物激素,广泛参与细胞分裂、分化和器官发育,其信号转导过程高度复杂且精细,但同时也伴随随机性变化。本文结合最新研究成果,深入解析拟南芥花药分生组织中生长素介导的随机基因表达特征及其生物学意义。 拟南芥花药分生组织是一个多细胞组成的组织,其形态和发育过程经过严格调控。不同于单细胞系统,分生组织中基因表达不仅受细胞内分子噪声的影响,还受细胞间通讯及发育阶段的调控。
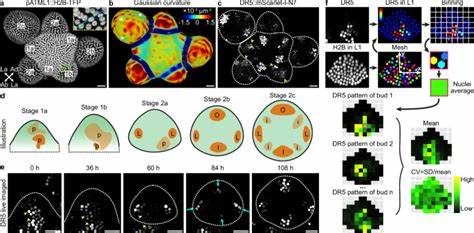
生长素信号经典传导机制为:生长素通过PIN蛋白的极性转运形成局部最大值,这些局部高浓度区域驱动器官原基的形成。细胞内,生长素结合受体TIR1/AFB促进Aux/IAA蛋白降解,释放ARF转录因子,激活下游靶基因表达。例如人工合成的DR5启动子被广泛用于监测生长素响应基因活性,但近年来研究表明,DR5驱动的基因表达存在显著的随机性,且该随机性主要源自转录和翻译过程中的内在噪声。 通过高分辨率共聚焦成像,研究人员观察到年轻花药分生组织阶段(1期至2a期),DR5表达呈现高度不规则的细胞斑点分布,这些斑点在不同花药之间位点不固定、大小和形状多变,反映出基因表达的显著随机性。随着发育进程推进至2b至2c期,这些随机斑点逐渐整合并形成四个明确定位的生长素响应高峰,对应未来的花萼原基位置。该动力学变化揭示早期高随机性的表达经过空间平均效应实现了整体信号模式的稳健。
针对随机性起源的探讨,研究排除了极性生长素转运和生长素本身水平的显著贡献。通过应用2,4-二氯苯氧乙酸(2,4-D)这种不受极性输运影响的生长素类似物,发现虽然整体生长素水平被提升,DR5表达仍呈现高度随机的斑点分布;同时,利用R2D2报告系统监测Aux/IAA蛋白降解的生长素感知,发现其空间分布模式高度一致且稳定,显示感知层面随机性较低。综上,随机性主要由基因表达的内在噪声驱动,即转录起始、染色质构象及mRNA降解等步骤中固有的分子波动。 为定量评估基因表达中的噪声,研究利用双荧光报告系统,分别驱动两个不同颜色荧光蛋白表达于同一启动子下。该系统允许区分由细胞外环境和细胞内部分子噪声引发的表达变异。结果表明,内在噪声对DR5启动子驱动表达具有支配性,在同一细胞内两个报告基因的表达水平高度不一致,证明基因表达存在显著的随机成分。
此外,该内在噪声在发育过程中基本保持稳定,体现了随机性作为发育过程中的固有属性。 在研究范围内,不仅是DR5,拟南芥内源性生长素响应基因如AHP6和DOF5.8也显示出随机基因表达特征。然而,相比DR5,这些基因的表达噪声幅度较低,并表现出更明显的空间与时间依赖性,表明内源基因通过更复杂的调控机制部分抑制了随机性。这可能与其启动子区域结构复杂、多样的转录因子结合位点及调控元件有关,使得表达更加稳健,确保关键发育过程的精准推进。 值得注意的是,尽管细胞层面表达噪声持续存在,整体组织水平的生长素响应模式却随着细胞数目增加而表现出更高的稳健性。初期分生组织细胞数目较少,随机表达造成的异质性难以被平均,导致整体信号模式不稳定。
随着器官发育,细胞大量增殖,随机表达信号在空间上实现了“平均效应”或“空间平滑”,使得局部的随机波动被抵消,最终形成稳定且可重复的生长素信号空间模式。实验证据显示,阻断细胞分裂(如通过药物奥立扎林处理)减少分生组织细胞数,会显著提高全球信号模式的随机性,支持上述空间平均假说。 该研究不仅深化了对植物发育过程中随机基因表达的理解,还揭示了植物如何在充满分子噪声的环境中维持发育稳健性。它表明基因表达的随机性并非单纯的干扰因素,而可能被植物发育系统灵活利用,通过多重尺度的调控机制实现组织功能的鲁棒性。同时,它提示发展生物学研究需关注基因表达的多样性及其空间拓扑分布,结合细胞数量和组织形态学变化,综合解析发育过程的复杂性。 未来,进一步探索不同基因启动子的结构特征及其对随机表达调控的影响,将有助于揭示基因表达随机性的本质机制。
此外,解码基因表达随机性与表观遗传调控、细胞周期状态以及细胞间通讯之间的相互作用,将推动植物发育调控网络的全景式理解。该领域研究成果亦将为作物遗传改良和植物组织工程提供理论基础,促进农业与生物技术的创新发展。