大麦作为人类最早驯化的谷物之一,其驯化与传播历史长期为考古学家和遗传学家所关注。随着全基因组测序和古DNA技术的发展,研究者不再只依赖少量标记或单基因分析,而开始从单倍型(haplotype)层面揭示大麦基因组的拼接历史。最近发表于Nature的研究以单倍型为核心,通过对现代与古代大麦全基因组的系统分析,提出了关于驯化起源、基因渗入与后驯化分化的全景式图像,为理解大麦如何成为全球作物提供了重要的基因组证据。 研究背景与思路 传统上,许多作物的驯化研究曾试图寻找单一中心,但基因组数据不断提示一个更加复杂的面貌。大麦的驯化并非单点触发,而是体现为多个野生群体对栽培种基因库的层层贡献。研究团队采用单倍型为单位,而非单个位点或染色体整体来追踪遗传物质的起源。
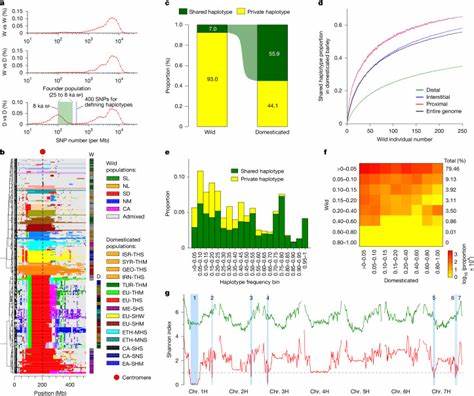
这种方法强调在基因组局部尺度上确定哪些片段源自哪些野生种群,从而重建驯化群体是如何由不同野生来源"拼接"而成的。 样本与方法概述 研究整合了682份基因库样本的全基因组序列和23份考古时期的大麦谷粒古DNA。野生大麦样本覆盖了西亚不同地理区系,将其分为五个主要野外种群,分别分布于南黎凡特、叙利亚沙漠、北黎凡特、北美索不达米亚和中亚。作者使用IntroBlocker等工具定义祖先单倍型群(AHG),并结合PSMC等人口历史推断方法和古DNA数据,追溯单倍型的时空来源与进入栽培群体的时间窗。 关键发现:马赛克式起源与主导区域 研究显示,现代栽培大麦的基因组呈明显的马赛克结构,长片段的单倍型在近端(靠近着丝点的低重组区)与远端有不同的多样性格局。大约56%的栽培单倍型能在至少一个野生样本中找到对应者,表明广泛的野生来源参与了栽培群体的形成。
以时间划分,约91%的可追溯单倍型在32千年至8千年前这一阶段分化并进入栽培基因库,这意味着驯化前及早期栽培阶段的野生多样性为后来的栽培群体提供了绝大部分"原料"。不同地理野生群体对栽培大麦的贡献存在差异,南北黎凡特与中亚的野生群体各自贡献约20%至27%,而叙利亚沙漠和北美索不达米亚的贡献相对较低但仍不可忽视。 单基因镜像:关键驯化基因的起源与年龄 将视角聚焦到几个决定性驯化性状的基因,研究对BRITTLE RACHIS(Btr1/2)、SIX-ROWED SPIKE(Vrs1)和NUDUM(Nud)等基因周围的单倍型进行了年代估算与谱系溯源。结果表明,不同的驯化相关突变多次独立发生且分布于不同野生种群。例如非脱落穗(非脆轴)相关的btr1单倍型在大约2.7万年前出现,btr2约1.5万年前出现,六行穗相关的某些等位基因最早可追溯到2.5万年前,而nud导致裸粒性的突变约在1.6万年前出现。这些年代普遍早于或接近考古学上观察到的栽培形态出现时间,暗示某些"驯化性状"在被人类选育固定之前可能已在野生种群中以低频存在。
古DNA证据:驯化基因组的持久结构 古DNA样本覆盖公元前6000年至公元前2000年间的多处以色列遗址。高质量古基因组显示,古代大麦与现代栽培型在基因组多处保有相同的长片段单倍型,尤其是在近端低重组区。这一发现说明,早期农耕者并非从某一当地野生群体直接驯化而来,而是使用了一个更为广泛的、已含来自多方野生源的"创始"栽培群体。古代样本的基因多样性呈现时间变化的梯度,晚近的样品显示较强的西方来源基因流,反映古代贸易与人群移动对农作物基因成分的影响。 人口史与地理扩散 通过基于单倍型的分歧时间估算,研究揭示出栽培大麦的主要分化事件与农耕传播路径时间点相吻合。约8500年前,近东和欧洲、东亚与埃塞俄比亚方向的栽培大麦群体开始分化,随后在7500年前欧洲内部出现了两个主要谱系(两行与六行为主),并在7千至5千年前完成更细分的区域分化。
随着向外传播,外部群体与当地野生种群之间的基因流逐渐减少,区域性谱系通过地理隔离与农业实践差异逐步固定独特的基因组拼贴。 基因组学与考古学的互补价值 研究的一个重要启示是基因组数据如何补充并解释考古学证据。比如在某些旧石器或新石器遗址发现的早期栽培型形态可能反映出一种"原始耕作"或"前驯化"状态,其中关键性状尚未完全固定。单倍型层面的证据允许科学家追溯这些形态在更长时间尺度上的出现与流散,为建立考古时间线提供分子钟辅助。同时古DNA揭示的区域间基因流有助于解释为什么在考古层序中会出现突如其来的作物形态变化。 对育种与遗传资源保护的启示 从实用角度看,高度的单倍型差异性既是机遇也是挑战。
多样的野生来源意味着现代育种可以从不同的野生群体中挖掘抗逆性、病害抗性或环境适应性相关的有利片段。然而,深度分化的单倍型结构也会妨碍基于关联分析的性状定位,当不同群体之间完全不共享某些单倍型时,传统的群体对比方法很难区分选择信号与祖先结构的影响。因此,育种研究需要结合突变组学、基因功能验证与多来源联用策略,以便在复杂的基因组背景中精确识别因果位点。保护方面,研究表明野生大麦的不同地理群体对栽培基因库均有贡献,因而维护野生种群多样性对未来作物改良至关重要。 方法学反思与局限 尽管单倍型基础的研究提供了前所未有的分辨率,但也存在方法学与解释上的局限。分子钟估计受限于突变率的准确性与自交作物的遗传特性,年代值应以相对顺序为主而非绝对精确年表。
此外,古DNA样本数量与地域覆盖尚不足以完全重建每一步基因流事件的细节。某些长时间保留的近端单倍型可能受到选择或结构变异(如倒位)的影响,需结合更多的图谱化长读与结构变异分析来澄清。 未来方向 后续研究可在多个方面展开。扩大古DNA地理与时间跨度将更好地揭示迁移与贸易对基因组构成的作用。结合高质量全基因组拼接、pangenome框架与功能组学,将有助于解读结构变异与基因表达在驯化与环境适应中的角色。育种上,基于单倍型的"祖先片段"筛选方法可用于有目的地引入野生有利单倍型,同时避免不良基因背景。
保护政策应更积极地将野生大麦群体作为遗传资源的重要来源,建立区域性保护与活体材料库,支持未来应对气候变化与病虫害压力的育种需求。 结语 从单倍型视角解读的大麦驯化历史揭示了一个复杂而渐进的过程:野生群体的多重贡献、早期的前驯化阶段、长时间的基因流与最后的地理性分化共同塑造了现代大麦的基因组马赛克。此类研究不仅丰富了对人类农业起源的科学叙述,也为现代育种与遗传资源管理提供了重要的战略信息。未来在更广泛的时间与空间尺度上整合古DNA、全基因组与功能验证,将有望把大麦这一古老农作物的演化故事讲得更加清晰与完整。 。