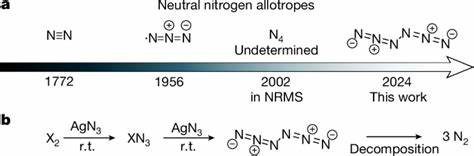
基因组结构的变化是生命演化的核心驱动力之一,而最近的研究发现,非海洋环节动物中的基因组发生了一次间歇性的、规模空前的重排事件。这种现象不仅打破了传统认为基因组结构稳态主导演化的观点,也为生物多样性产生和进化机理提供了重要案例。本文将深入解析这种基因组重排爆发的特征、成因以及其对环节动物进化的深远影响。环节动物是一个多样化的动物门类,涵盖了海洋、淡水和陆生环境的众多物种。虽然海洋环节动物的基因组结构通常保持高度保守,但非海洋环节动物如泥鳅类和蚯蚓类,在基因组水平表现出惊人的重排频率和幅度。这种基因组“重洗牌”现象表现为基因位置的大范围迁移、染色体结构的突破性变化以及调控网络的重新构建。
研究者通过全基因组测序和染色体重构技术,揭示了非海洋环节动物基因组广泛的线状序列扰乱,特别是键基因群如Hox基因簇的重新组织。Hox基因对于体节模式和发育过程至关重要,传统上其在基因组中的局部紧密排列对其功能保守性起到保障作用。然而,这些环节动物中长距离相互作用的新形成表明,基因组三维结构的塑性能够为功能调控提供新的空间框架。基因组重排的诱因与驱动机制成为科学界极为关注的话题。当前研究指出,这些大规模重排事件与部分关键基因的丢失有关,例如与基因组稳定性及细胞分裂稳态密切相关的基因的损失,可能引发细胞内基因组重构机制的失控。此外,跨转座元件的活跃插入导致了新型假着丝点(neocentromeres)的形成,促进染色体片段的重组和非典型染色体的出现。
转座元件的作用不仅仅在于破坏基因组稳定性,同时也为新基因产生和调控网络创新创造了原材料。与基因组重排并行,非海洋环节动物基因组经历了复杂的基因家族演化和功能重塑。大量新兴基因族谱的出现,且部分基因表达在应激反应和适应性进化中获得显著差异,使得物种能够更灵活地适应内陆淡水和陆地复杂多变的生态环境。值得强调的是,非海洋环节动物显示出一种不同于其他动物门类的基因组结构演化宽容性。尽管多数动物门类基因组保持高程度的宏观共线性以维持基本功能和发育稳态,这些环节动物则表现出极强的染色体重构容忍性,反映出基因组演化过程的独特模式。这一发现深刻挑战了我们对基因组稳定性与进化创新关系的传统认知。
此类基因组变革通常伴随着物种群体历史上的快速分化阶段,也就是生物学中所谓的“间歇性爆发”。这种爆发结束了一个长期进化相对稳定的阶段,标志着新物种阵营的产生和新适应性特征的形成。研究表明,非海洋环节动物的基因组重排过程与其向淡水和陆地生态系统的转移密切相关,这不仅改变了其生态位,也促进了生理和形态特征的多样化。基因组结构的重塑同样影响了染色体的三维空间组织。通过高级的染色体构象捕获技术(如Hi-C),研究者观察到重排造成的局部和远程基因调控联系被重新连接,形成了新的拓扑关联域。这说明基因组三维构象的重塑是基因表达调控和发育过程适应环境变化的重要基础,体现了立体基因组组织对结构和功能演化的双重影响。
此外,间歇性的大规模基因组重排与染色体不稳定性有关,这种现象在癌症细胞和其他高变异生物体中也有体现。研究非海洋环节动物基因组重塑为理解更广泛的基因组塑性机制提供了宝贵窗口,也为研究染色体结构异常及其功能后果带来启示。转座元件的激活和新假着丝点的形成,不仅为染色体创新提供动力,还体现了基因组中“冲突与创新”的动态平衡。整体而言,这些研究成果显示了基因组演化的高度动态性和复杂性,表明生命进化不仅仅是基因序列的缓慢积累,更包含着结构层面的剧烈变化和调控层面的深度再造。非海洋环节动物基因组中发现的间歇性重排爆发现象,不仅丰富了我们对基因组进化的理解,也为研究生物适应性策略提供了新的思路。通过未来更多基因组数据积累、多物种比较分析以及功能验证实验,我们有望解锁这一现象背后的分子机制和生态进化意义。
综上,非海洋环节动物经历的基因组间歇性爆发式重排事件展示了基因组高度可塑性与生态适应之间的紧密联系,揭示了复杂生物进化历程中的一段关键篇章。