基因表达的随机性是生命系统中普遍存在且极具意义的现象。植物作为多细胞生物,其发育过程中基因表达的时空异质性在细胞命运决定和器官形成中发挥着关键作用。拟南芥作为模式植物,在发育生物学研究中具有重要地位。近年来,围绕拟南芥花蕾中生长素信号传导所驱动的基因表达随机性展开的研究,揭示了细胞内分子噪音对基因表达调控的深刻影响,为理解植物发育的稳健性机制提供了新的视角。生长素是调节植物发育的核心激素,其通过介导细胞间的信号传递形成功能性模式,指导器官的发育与分化。在拟南芥花蕾分生组织中,生长素的定位和响应被视为花器官起始的关键调控节点,而基因表达的随机性是否存在以及如何影响发育进程,长期以来一直是学术界关注的焦点。
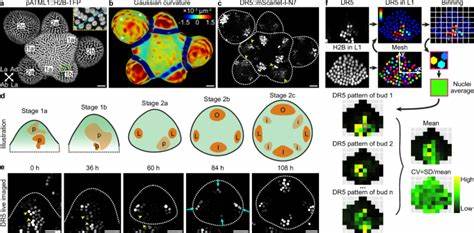
通过引入DR5这一人工设计的生长素响应元件回报系统,研究者首次对生长素响应基因在花蕾中的表达模式展开了定量分析。令人惊讶的是,在发育早期的花蕾中,DR5表达呈现出极高的变异性,细胞水平的表达存在随机的高表达斑块,这种分布在不同花蕾之间差异显著,显示出生长素信号响应的时空随机性。伴随着花蕾发育进程进入成熟阶段,这种随机性逐渐缓和,表达模式趋向于稳定,形成四个明确定位的生长素响应高峰,对应即将形成的萼片原基区域。解析该现象的关键在于确定随机性的根源。通过过量给予2,4-D(生长素类似物)以削弱极性输送对内源生长素分布的影响,发现细胞间信号差异虽然有所降低,但仍未消除表达模式中的随机性。此外,通过R2D2系统反映的生长素感应强度数据显示,生长素感受过程本身具有相对稳定的空间分布,变异程度远低于DR5表达,表明随机性主要源于下游基因转录调控过程中的内在分子噪声。
为更精细区分内源噪声与外源噪声对基因表达波动的贡献,研究者构建了基于双荧光蛋白标记的DR5双报告系统,允许同时监测同一细胞内两个独立的DR5驱动报告的表达。结果显示,两个信号之间存在显著的不相关性,直接证明内在分子噪声是生长素响应基因表达随机性的主要成因,且这一随机性贯穿整个花蕾发育早期至成熟过程。相比之下,来自内源生长素信号通路下游的天然靶基因AHP6和DOF5.8表现出更低的随机基因表达水平,其噪声幅度和时空分布具有一定的特异性和调控。这或许体现了进化层面对关键调控基因表达的精细调节,以维持发育的稳健性和准确性。值得关注的是,即使细胞水平的基因表达随机性持续存在,整体组织尺度的生长素响应模式依然表现出高度的稳定性和可重复性。对此,研究提出了“空间平均”理论,即随着花蕾发育和细胞数量增加,细胞间的随机表达经过空间整合与平均作用,从而实现组织层次上的模式稳健。
通过药物如奥立铣和羟基尿嘧啶抑制细胞分裂,减少花蕾中细胞总数,实验观察到整体生长素响应模式的变异性显著上升,但单细胞层面的噪音水平未见明显变化,进一步支持了空间平均机制的有效性。对此理解,不仅丰富了植物激素信号调控网络的内涵,也为解释植物如何在分子噪声存在的情况下,保证形态发生的高度精细和一致性提供了理论基础。从分子机制角度看,不同基因启动子结构对噪声水平也具调节作用。DR5启动子包含多串紧密排布的高亲和力生长素响应元件,可能因其结构允许ARF转录因子高效结合而产生较高的转录偶发性,从而导致较大噪声。相比之下,AHP6和DOF5.8的启动子拥有较少且间隔较远的响应元件,并引入多样化的结合位点以及辅助结合元件,这些因素有助于提升转录调控的特异性和稳定性,减少随机波动。这一发现提示,启动子设计和转录因子结合特性是基因表达噪声调节的重要层面。
进一步结合转录后调控,表观遗传修饰以及转录因子复合物动态,未来有望全面揭示基因表达随机性产生及其生物学意义。在植物发育与环境适应之间,基因表达噪声既是一把双刃剑。其随机特性通过产生细胞异质性,为组织的灵活发育和对环境变化的敏感性提供潜在基础,有利于塑造多样化的细胞命运和组织形态。与此同时,噪声需被有效调控以避免干扰正常发育过程,保证遗传程序的可靠执行。拟南芥花蕾中生长素响应基因表达随机性的研究为此提供了生动范例,揭示了噪声生成机制与缓冲机制的共存与协调。整体来看,通过构建和分析基于多维度荧光标记的活体成像技术,研究者能够定量捕捉拟南芥花蕾中生长素信号路径的动态表达变化,揭示分子噪声对基因表达的深刻影响及组织尺度上的稳健调控。
该研究不仅深化了对植物激素信号和发育调控的理解,也为多细胞有机体中随机基因表达的调节机制研究开辟了新方向。未来,结合系统生物学、单细胞测序及多组学技术,有望进一步解析基因表达随机性在植物发育、环境响应与表型塑性中的多层次作用,推动植物科学和农业创新发展。