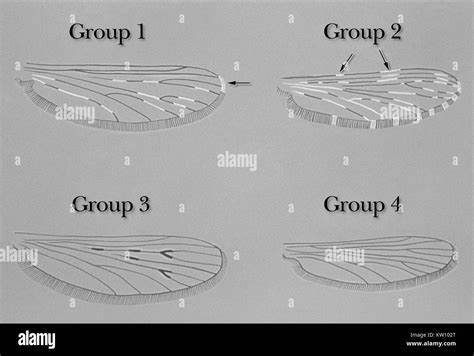
在生物学领域,纵观进化的长河,微观进化与宏观进化之间的联系一直是科学研究的焦点。微观进化通常指的是种群内部基因频率的变化,是短期内适应环境的过程;宏观进化则是指物种之间的大尺度分化和演化,通常涵盖数百万年的时间跨度。长期以来,科学界关于这两者之间是否存在内在联系,甚至微观进化能否真正驱动宏观进化,存在诸多争议和不同观点。最新的研究通过对果蝇(Drosophila spp.)翼形态多层次的变异分析,为这一问题带来了崭新的视角和证据。 果蝇翅膀作为形态学研究的经典模型,因其结构复杂且易于测量,成为探讨发育和进化机制的理想对象。研究团队通过分析覆盖六个生物层级的果蝇翼形变异数据,从个体内的发育噪音到跨越四千万年的物种宏观进化,全面揭示了微观与宏观变异之间的正相关关系。
这一发现颠覆了传统认知中,发育噪音与宏观进化之间关联较低的假设,强调了发展系统和长期适应性峰值波动历史之间的相互作用。 研究的核心在于检视不同层面上的变异及其与进化分化的关系。从最细微的发育波动(发育噪音)到种群内的基因变异(遗传变异),再到种间的形态多样性,这些变异通过统计学上的协方差矩阵进行量化和比较。结果令人惊讶地显示,发育噪音与宏观进化速率之间的相关度远超遗传变异与种群分化的程度。换言之,个体发育过程中产生的细微不确定性不仅反映了组织结构的稳定性与变异潜能,也与长时间尺度上物种翼形的演化模式紧密相联。 这种跨尺度的变异连结支持了一种被称为“内在一致性假说”(congruence hypothesis)的理论框架。
该假说认为,一个生物体的发育系统能够反映环境和选择的长期动态,它的变异性并非单纯由当前遗传变异驱动,而是受累积的宏观进化历史塑造。换句话说,微观层次的变异性实际上是宏观进化过程的镜像,表明发展过程在进化中的调节作用被低估了。 与此相对的是传统的“遗传约束假说”,该假说认为遗传变异矩阵直接限制并引导种群分化的方向与速度。研究中的数据表明,尽管遗传变异(G矩阵)与种群分化(Dpop矩阵)存在明显正相关,但其相关性与发育噪音(F矩阵)和宏观进化速率(R矩阵)的相关程度相比,显得较弱且不够充分说明全部演化过程。 通过细致的统计分析,研究团队发现,发育噪音与突变变异之间也存在强烈的正相关,这表明不同类型的表型变异虽来自不同来源,却可能共享发育系统这一共同的调控机制。这种机制使得个体在面对基因和环境扰动时表现出相似的变异模式,使微观变异能够以某种方式映射至宏观演化轨迹。
除变异关系外,研究还探讨了形态发育中的异速生长(allometry)对这一现象的潜在影响。虽然异速生长是果蝇翅膀大小与形态变化的重要因素,且不同部位的生长比例差异可能影响整体形态的变异模式,但先前与本研究的结果均显示,消除异速生长因素后,变异矩阵之间的强烈关联依然存在。这说明形态变异的内在一致性不仅仅是由尺寸调控机制造成,而是更深层次的发育与进化系统属性的体现。 在方法上,此项跨尺度研究运用了多元统计模型和贝叶斯混合效应模型,结合了各种数据采集技术,包括基因来源的遗传变异测定、不同环境处理下的表型塑性评估以及个体翅膀两侧的微小差异计算。这些方法的综合运用确保了对变异结构的准确估计与层级间关系的科学验证。 该研究的意义深远,它不单纯为演化生物学中的一个理论争议提供了数据支撑,更是促进了跨层级、生物系统动态理解的发展。
它强调了观察和解释生物变异时,不能仅局限于遗传层面的线性思维,而应考虑发育系统、环境和进化史三者的复杂交织。特别是在理解物种适应性和多样性生成的机制上,提出了基于发育变异和历史适应动态的多因果解释框架。 未来的研究有望基于这一理论框架,进一步探讨各类生物性状的跨层级变异机制,特别是在快速环境变化下种群如何利用发育噪音及其产生的变异应对挑战,以及这种机制是否普遍适用于其他物种甚至更广泛的生物系统。同时,发展数学模型和计算工具以量化不同进化力对变异及分化的综合影响,将促进演化理论的整合与创新。 总之,果蝇翅膀形态的跨尺度变异研究揭示了微观进化与宏观进化之间深刻且复杂的内在联系,表明发展系统不仅塑造个体的变异潜能,也在宏观进化史上留下不可抹去的印记。这一发现推动了理解生物多样性生成的核心问题,为构建涵盖不同时间和空间尺度的统一进化理论奠定了基础。
。