多细胞生物体的成功进化深受其增大体积的影响,与单细胞祖先相比,多细胞体型的增大带来了多种生态和功能优势。然而,随着生物体的体积增大,营养物质的运输问题成为限制其进一步成长的关键瓶颈。传统观点认为,大型多细胞生物必须依赖专门的运输机构,如血管系统或纤毛运动,实现有效的物质输送,否则仅靠扩散过程难以满足内部细胞的营养需求,导致生长放缓甚至停滞。近日,通过对实验进化的雪花酵母群体的深入研究,科学家们发现了一种新颖的代谢驱动流体流动机制,有效突破了扩散极限,使多细胞酵母簇得以在宏观尺度上保持指数级的快速增长。 雪花酵母是利用酿酒酵母(Saccharomyces cerevisiae)通过不完全细胞分裂形成稳定的细胞链条集群,这些细胞链以机械缠结的方式形成坚固的大型群落。令人惊讶的是,这些群落直径可达毫米级,远超过传统理论所预测的通过扩散满足细胞内部营养需求的阈值。
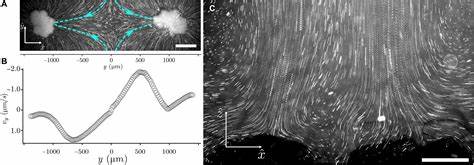
更为重要的是,雪花酵母并未进化出任何典型的多细胞营养运输适应结构,如血管或纤毛,因此其实现大规模生长的机制长期以来备受质疑。 最新实验显示,当这些大型雪花酵母簇悬浮于液体培养基中时,簇内细胞的代谢活动会促使其周围流体产生大尺度的循环流动。具体来说,流体从群体侧面流入,经过簇体内部,并从顶部流出,形成持续的流动通道和营养循环体系。这种流速与某些拥有纤毛或鞭毛的多细胞生物如沃尔沃克斯藻等相当,彰显了这一物理机制在无结构辅助下的高效能。此外,这种代谢产生的流体流动不仅增强了营养物质的运输效率,还使整个群体能够维持一致、稳定的营养供给,实现整个群体的指数级细胞分裂和增长。 更深入的研究中,通过对流体微粒追踪实验和粒子均方位移分析,科学家们发现流动的动力源于代谢引起的流体密度梯度。
酵母细胞在发酵过程中会消耗培养基中较重的葡萄糖,产生密度较低的乙醇和二氧化碳,导致局部流体的密度降低,进而驱动浮力差异产生的流体环流。实验中,将培养装置倒置后流动方向亦随之反转,有力证实了流动机制与重力方向密切相关,由代谢产物介导的流体密度驱动浮力流动成为主导动力。此外,实验还排除了其他可能性,比如表面张力差异导致的马兰戈尼流动或蒸发驱动的对流,确保流动的唯一来源为代谢活动引发的密度梯度。 关键的是,这种流动机制只在簇达到一定体积阈值后才显现。早期进化阶段较小的簇体因整体代谢产物不足以形成显著密度梯度,流体环境依旧靠扩散占主导,无法形成明显的流动。随着实验进化的进行,簇大小逐渐增大,代谢总量积累到足以驱动宏观浮力流,流体运动表现出明确的加速化趋势。
当大型簇断裂成小簇时,流动现象随之消失;相反,通过连接多个小簇形成更大结构,流动又重新出现,这进一步表明簇体规模对于流动机制至关重要。 生物物理学意义上,酵母代谢驱动的流动机制为多细胞有机体克服营养运输的物理瓶颈提供了全新思路。在传统理论中,体积增大常被看作营养扩散效率递减的死胡同,多细胞生物必须通过演化血管或纤毛等运输系统方能持续增长和复杂化。此类结构需要复杂的基因调控和形态学创新,且进化过程耗时漫长。代谢驱动的流动机制则直接依赖于代谢活动本身,无需特殊结构或基因指导的创新,即可实现功能的跃迁,成为一种“生物物理支架”,为多细胞生物的快速增长提供基础。它为我们理解多细胞演化早期的机制打开了崭新的视野,也说明简单物理法则在生物进化中拥有极大塑造力。
此外,雪花酵母群体的多孔结构和细胞链缠结特性为流体流动提供了理想的通道。与某些细菌生物膜或群体因胞外基质密集阻滞流体不同,雪花酵母的结构使外部流体能够深入簇体内部,形成有效循环,有助于营养和废物在内部快速交换。这种结构与生理功能的高度耦合为多细胞体积巨大化提供了新的微观机制解释。 从生态和进化的角度看,代谢驱动流体流动不仅促进了群体内部的均匀营养供给,也为避免因资源不均诱发的细胞死亡或停滞奠定基础。维持指数级的增长速度,有助于群体快速占领生态位,提升竞争优势。未来该机制或能在其他原始多细胞或群体生活的微生物群落中被发现和应用,提示演化中生物与物理环境相互作用的普遍性。
这一发现对现代生物工程和应用微生物学亦有深远启示。理解和利用代谢驱动的流动可以设计创新的发酵系统和组织培养平台,促进细胞或微生物团体的高效生长。借助物理原理调控流体环境,为工业生产带来节能高效的新思路。此外,此机制的发现可能推动人工多细胞系统的开发,激发新型生物材料和合成生物学技术的进步。 综上所述,代谢驱动的流体流动机制成功突破传统扩散限制,成为大型多细胞酵母群体实现宏观规模指数级生长的关键推动力。这种物理与生物相互作用的涌现现象,不仅深化了我们对多细胞起源和演化的理解,也为未来生物学与工程学交叉领域提供了丰富启发。
世界各地的研究者正积极探索其更广泛的生物学意义及潜在应用前景,期待在解码生命复杂性的道路上继续前行。









