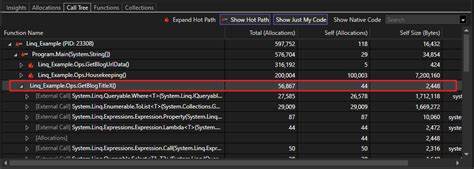
RNA并非只是从DNA到蛋白质之间的简单信息中介。近年来,越来越多的研究表明,RNA具有惊人的结构多样性与功能可塑性,而其中"非典型分子构象"(unusual molecular conformation)正在成为解释RNA多功能性的关键线索。非典型分子构象是指RNA在自然条件或特定环境下采用的不同于常规二级结构模型的折叠形态,包括不规则的干涉结构、稀有的碱基配对方式、及复杂的三级相互作用。这些特殊构象不仅赋予RNA新的识别与催化能力,还为调控网络和进化创新提供了物理基础。理解这些构象如何产生、如何被识别以及如何影响功能,对于基础生物学和转化医学都具有重要意义。RNA的结构层级从一级序列到二级结构再到三级折叠,是决定其功能的根基。
传统观点强调经典的碱基配对和干涉螺旋,例如A-U、G-C配对以及发夹、茎环等局部结构。然而,随着高分辨率成像技术和单分子检测手段的发展,科学家发现许多RNA分子能够形成不符合常规配对规则的结构,例如错配碱基配对、三联体构象、G-四联体以及反常的二级结构拓扑。这些非典型构象常常出现在功能中心或调控位点,显著改变RNA与蛋白质、小分子或其他核酸的相互作用方式,从而扩展其生物学角色。识别和解析这些非典型构象依赖于多种实验与计算方法的结合。X射线晶体学长期以来为解析高分辨率的RNA结构提供了重要数据,但RNA结晶困难,且动态构象常被结晶条件所固定。核磁共振(NMR)可以捕捉溶液中RNA的动态信息,适用于中小型RNA体系,揭示瞬态构象和短寿命中间态。
近年来兴起的冷冻电镜(cryo-EM)则在解析大型RNA及RNA-蛋白复合体方面显示出强大优势,能够观察到异构混合态。单分子荧光共振能量转移(smFRET)和化学探针测序(如SHAPE-seq)等技术能够提供动力学与构象群体信息。与此同时,计算模拟,尤其是基于原子级分子动力学(MD)和机器学习的结构预测方法,为捕捉短寿命但功能关键的非典型构象提供了必要补充。多方法整合已成为研究RNA非典型构象的主流策略。非典型构象在RNA功能中的角色可以从多个经典例子中得到说明。核糖开关(riboswitch)是调控代谢通路的天然传感器,通过结合小分子诱饵改变RNA构象以开关基因表达。
很多核糖开关展示出非常规配对及三级相互作用,其非典型构象使其对配体的识别既高度专一又能实现大幅构象转换。核糖酶(ribozyme)是RNA催化的代表,一些具有催化活性的RNA通过错配、碱基堆叠的畸变以及金属离子介导的非典型构型形成活性位点,从而实现化学转换。剪接体中的小核RNA(snRNA)与前体mRNA的复杂折叠也涉及短暂且罕见的构象,正是这些构象允许精确识别剪接位点并完成剪接反应。病毒RNA经常采用G-四联体或伪节等稳定但不常见的三维结构,用以调控翻译、复制和逃避免疫监视。非典型分子构象的出现有多重原因,包括序列本身的内在倾向、离子环境(尤其是镁离子等二价阳离子)的影响、与蛋白质或小分子的结合驱动,以及转录过程中瞬时的共折叠动力学。RNA在细胞内不是孤立存在的,其折叠路径常受伴随分子(伴侣蛋白、核糖体、翻译因子等)和细胞器环境影响。
共转录折叠意味着RNA在合成过程中可能被"捕获"进某些非典型构象,进而影响后续功能。环境变化如pH、温度、金属离子浓度或配体存在也能诱导构象断裂或重折叠,从而实现快速响应。这些因素共同塑造了RNA的构象能量景观,使得同一序列能够在不同条件下切换到不同的功能状态,体现出高度可塑性。理解非典型构象对RNA演化的贡献同样重要。RNA世界假说提出,早期生命可能极度依赖RNA既作为遗传信息载体又作为催化分子。非典型构象提供了额外的功能维度,有利于在有限的序列空间中产生多样功能。
快速切换构象能力能够为原始分子提供可选择的功能阵列,使得少量序列能承担多重生物学任务,有助于在资源受限的早期地球环境中实现进化创新。此外,非典型构象增加了RNA适应突变和环境变化的灵活性,可能在分子进化过程中发挥桥梁作用,连接RNA功能与复杂蛋白质机器的演化。在医学与生物技术领域,非典型RNA构象的发现带来了新的靶点与工具。RNA分子构象的多样性意味着可以设计更精确的药物来靶向特定的非常规结构,例如针对G-四联体、伪节或独特的活性位点的小分子抑制剂或稳定剂。这一策略在抗病毒、抗肿瘤和调控代谢通路上显示出潜力。核酸适体(aptamer)的设计也可以利用非典型构象增强结合亲和力与选择性。
RNA疗法,包括siRNA、mRNA疫苗和反义寡核苷酸,其效果和稳定性在很大程度上取决于RNA的构象与被细胞识别的方式。通过工程化非典型构象,可显著提升RNA药物的体内稳定性、翻译效率和免疫原性调控。尽管前景广阔,但研究非典型构象仍面临挑战。首先是技术限制:许多非常规构象是短寿命或低丰度的,目前的成像方法可能无法在生理条件下同时提供高分辨率与动态信息。其次是解释复杂性:RNA构象能量景观高度多峰,单一测量结果往往难以唯一确定功能相关构象。数据整合与模型验证因此变得尤为重要。
再者,细胞环境的复杂性也使得体外研究结果难以直接外推到细胞内情境,需要发展体内成像与化学探针策略来捕捉真实的构象动态。最后,设计以非常规结构为靶点的药物仍需克服选择性和毒性问题,特别是当目标结构在不同RNA中存在保守性时,如何避免脱靶效应是必须解决的难题。未来研究方向可以从方法学和应用两条主线并进。一方面,推动多模态成像与测序技术的整合,开发能够同时获取高时间分辨率、高空间分辨率以及群体异质性信息的新型平台,将有助于描绘RNA构象切换的全景。机器学习与大规模模拟结合实验数据,可用于预测罕见但功能关键的构象并指导实验验证。化学生物学工具的创新,例如特异性标记、可逆交联和原位化学探针,将使研究者能够在细胞原位捕获非常规构象。
另一方面,在应用层面,将这些基本发现转化为药物开发、诊断工具和合成生物学部件的设计原则,是未来的重点。基于非典型构象的分子开关、传感器和催化元件能够丰富合成生物学的工具箱;而针对病原体或肿瘤中独特RNA构象的精确药物可能带来治疗学突破。总而言之,非典型分子构象为解释RNA多功能性提供了新的视角。这些特殊的折叠形态不仅扩展了RNA的化学与生物学能力,也为理解分子进化和开发新的生物技术提供了理论与实践基础。随着实验技术与计算方法的不断进步,我们有望更全面地捕获RNA的动态构象景观,并将这些知识转化为临床与工程应用。面对RNA研究的新时代,关注非典型构象及其生物学意义,将是揭示分子生命复杂性的关键一步。
。