植物作为地球上最重要的生物之一,其成长过程与细胞壁的力学特性密不可分。细胞壁不仅为植物细胞提供机械支撑,维持形态稳定,同时调节细胞的生长、伸展以及分化。近年来,科学家们发现植物细胞壁内部的纤维网络结构赋予了细胞壁极强的力学可调性,使其能够适应不同发育阶段的需求,保证植物正常发育。本文将深入探讨植物细胞壁中的纤维网络特性,以及其对细胞生长和整体植物形态塑造的意义。 植物细胞壁的主要成分包括纤维素微纤维、半纤维素和果胶类多糖。其中,纤维素微纤维是刚性较强的载重成分,通过与半纤维素和果胶构成的水合基质形成复合结构。
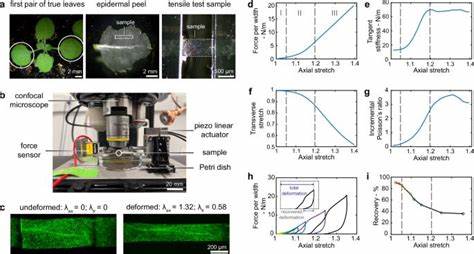
纤维素微纤维呈线状分布,构建成网络体系,这种网络结构决定了细胞壁的机械性质。纵观植物发育全过程,细胞壁需要在维持强度的同时具有一定的伸展性,以配合细胞膨压引发的扩张,保证细胞能够正常增长,同时避免过度破损。 探索植物细胞壁力学行为的一个有效途径是通过对阿拉伯芥叶表皮细胞进行张力测试。研究显示,表皮细胞壁呈现出三阶段的非线性力学响应。首先是在较低应变区,细胞壁比较柔软且韧性较好,有利于细胞初期的伸展;接着进入中间阶段,随着纤维素微纤维逐渐朝受力方向重新定向,材料表现出应变增强,使细胞壁刚度加大,抵御过度拉伸;最后进入高应变阶段,细胞壁刚度保持稳定,主要依赖于纤维素微纤维的拉伸变形,此时塑性变形开始显现,确保细胞壁在经历较大变形后不会破裂。不同开发阶段的细胞壁表现出不同的刚度水平,生长速度较慢的发育阶段刚度更高,这说明植物能够动态调节细胞壁的机械属性以适应不同的生长节律。
纤维网络结构是这种力学调控的关键。纤维素微纤维按照一定角度相互连接,形成类似菱形的网格结构,这种几何结构使得微纤维能够以弯曲、旋转和拉伸等多种方式协同变形。处于受力方向近乎垂直的纤维素微纤维初期主要通过弯曲使网络适应变形,随着负载的增大,更多微纤维重新定向后逐渐拉伸,导致整体刚度的增强。微纤维之间的连接体起到力的传递作用,同时连接强度的变化对机械性能的调控有显著影响。随着植物发育,连接强度增强,导致细胞壁整体刚度提升,更好地控制细胞生长速度和方向。 此外,研究中发现微纤维之间存在相对滑移现象,这使得细胞壁不仅体现弹性,还具备一定塑性。
通过加载-卸载循环测试,观察到其力学行为呈现出滞后和部分不可逆变形,反映了微观结构中的纤维之间可能出现的滑移和结构重组。这种塑性变形对于细胞在生长过程中调整形态具有重要意义,能够释放部分内应力,避免破坏性断裂,同时维持细胞壁的整体结构完整性。 细胞壁的力学各向异性也是植物形态生成的关键。纤维素微纤维主要沿特定方向排列,使得细胞壁在这一方向上的刚度明显高于垂直方向。这种各向异性决定了植物组织的生长方向和速率。例如spr2-2微管相关蛋白突变体表现出微管及其引导的纤维素排列更为有序,导致细胞壁机械性质明显各向异性,进而引起叶片的螺旋卷曲。
通过对不同拉伸方向的组织力学测试,得以确认纤维取向变化是导致器官形态改变的机械基础。 植物细胞壁力学研究不仅限于宏观组织层面,越来越多的技术实现了微尺度乃至纳米尺度的力学测量。例如利用共聚焦拉伸平台结合荧光微珠实现在线监测局部变形,结合有限元模拟准确区分材料本体力学特征与细胞结构的贡献。这些精细的测量揭示出细胞结构对整体组织力学影响较小,主要由细胞壁材料自身的纤维网络结构决定。这为细胞力学建模提供了坚实的实验依据。 力学模型方面,简单的仿射变形网络模型未能完全再现实验中观察到的三阶段非线性行为,表明只有重新定向纤维不足以解释细胞壁应变强化。
引入五梁模型,结合梁的弯曲、伸长和连接器的滑移,成功重现了实验观察的应变强化、变形各向异性及塑性特征。该模型不仅概念清晰,还能够解释材料属性随发育阶段调节的现象,突出连接器变形阻力作为调节力学性质的有效因子。 与动物组织力学如真皮的比较也带来了新的视角。真皮的胶原纤维网络与植物细胞壁的纤维素网络在力学表现上存在相似的非线性应变强化和明显的泊松比效应。然而,二者在纤维柔韧性及滑动机制上有显著差异,导致其机械响应细节不同。借鉴动物组织纤维网络的理论和实验结果,为理解植物细胞壁理论奠定了坚实基础。
探究植物发育中细胞壁纤维网络的力学可调机制,不仅深化了对植物生长和形态发生的理解,也为作物改良与生物材料设计提供了新思路。通过调节纤维素微纤维连接强度、排列方式及基质成分,植物实现了对细胞壁刚度和塑性的精准控制。未来,结合分子生物学和先进力学测试技术,将进一步揭示细胞壁结构与力学性能的动态调控机制,为植物科学和材料科学领域开拓更多创新应用。 综上所述,植物细胞壁中纤维素微纤维组成的纤维网络结构,凭借其独特的机械变形模式,实现了对细胞壁力学性质的高度可调。这样的结构允许植物细胞适应不同发育阶段的力学需求,保证细胞的安全延展和组织的稳定发育。探索和理解这一天然纤维网络的复杂性质,对推动植物生物力学研究和农业科技发展意义深远。
。









