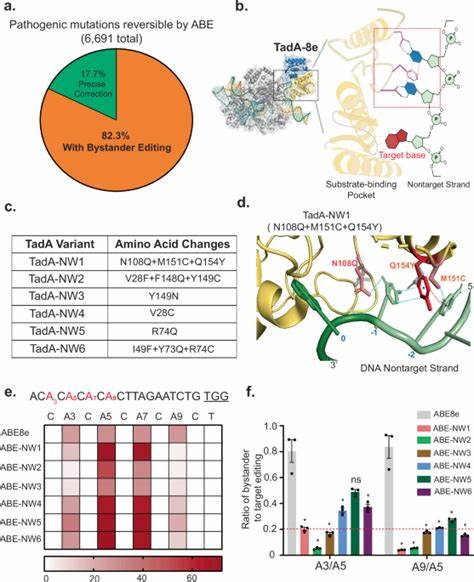
基因编辑技术,尤其是基于CRISPR-Cas系统的碱基编辑器,正迅速革新遗传病的治疗方法。碱基编辑器通过将特定的脱氨酶与失活的Cas9核酸酶融合,实现单碱基的精准转换,例如腺嘌呤(A)向鸟嘌呤(G)的转换,而无需产生双链断裂,极大地降低了潜在的细胞毒性和脱靶风险。然而,当前高活性的碱基编辑器如ABE8e虽然具有较高的编辑效率,却呈现出长达十个碱基对的宽泛编辑窗口,这使得在目标区域内多余的相邻可编辑碱基易被非特异性地修饰,形成所谓"旁观碱基编辑"问题。旁观编辑不仅降低了编辑精准度,还可能引起基因功能异常,进而带来严重的安全隐患,限制了碱基编辑技术在临床上的直接应用。对这一挑战的有效应对成为生物医学界的关键任务之一。 针对这一瓶颈,最近在Nature Communications上发表的一项研究提出了一种创新且简化的基于结构导向的碱基编辑器工程策略。
这一策略通过将天然的寡核苷酸结合模块巧妙地嵌入高活性的脱氨酶TadA-8e的催化活性中心,有效强化了对DNA非靶链的识别和结合能力,从而在保证目标碱基高效编辑的同时,将活性窗口缩窄至约4个核苷酸,显著降低了对邻近碱基的非特异性脱氨酶作用。该经改造的脱氨酶被命名为TadA-NW1,其与Cas9核酸酶结合后形成的ABE-NW1,不仅保持了与ABE8e相当的编辑效率,更在多个基因位点展示出优异的编辑特异性和大幅降低的脱靶活动。这种由结构生物学引导的蛋白质工程路径,有效避免了传统大规模定点突变筛选的繁琐步骤,极大地提高了编辑器的开发效率,展现出广泛的应用潜力。 机制上,TadA-NW1通过模拟人类Pumilio1蛋白的RNA结合结构域,建立起一系列新的碱基层叠作用、氢键及静电相互作用。通过这些特异性结合,TadA-NW1能够稳固DNA非靶链的U型构象,抑制脱氨基化反应的非靶向扩散,使脱氨酶更专注于欲编辑的目标碱基。这种稳定的结合模式不仅减少了编辑窗口的空间范围,同时降低了酶活性过快带来的过度编辑风险,最终实现了高效且精准的基因修饰。
这些改造不仅适用于活跃的CBEs(胞嘧啶碱基编辑器)和ABEs(腺嘌呤碱基编辑器),更拓展到新兴的碱基转化编辑器(例如腺嘌呤转化为胞嘧啶的ACBEs),研究表明对应包涵NW1突变的系列编辑器同样具备缩窄的活性窗口和提升的特异性,确证此结构工程策略的通用性和灵活性。这种多功能适应性为未来定制化碱基编辑器的开发奠定了基础,有助于匹配不同疾病相关基因突变和编辑需求。 临床验证方面,研究者特别挑选了具有高度临床需求的囊性纤维化(CF)治疗模型,聚焦CFTR W1282X突变的精准修复。此突变因AT含量丰富,传统碱基编辑器编辑该位点时往往出现严重的旁观编辑,导致不良氨基酸替换,影响蛋白功能恢复。通过体外在人肺上皮细胞中应用ABE-NW1联合针对性设计的sgRNA,成功实现了超过三分之一的完全矫正(无旁观编辑)的理想编辑产物,且显著提升了CFTR功能蛋白的表达水平。与ABE8e对照组相比,ABE-NW1修复效果更稳定,旁观碱基编辑显著降低,极大增强了治疗的安全性和有效性前景。
此外,该策略同样显著提升了编辑器的脱靶安全性。通过针对已知Cas9相关及非相关的DNA脱靶位点的检测,结果显示ABE-NW1在保持高编辑效率的同时,大幅降低了非目标位点的A-to-G替换率。整基因组测序亦未检测出显著新增单核苷酸变异,表明通过蛋白质工程引入的寡核苷酸结合模块并未产生不可控的脱靶效应,确保其临床应用的可行性和安全性。 从技术层面来看,这一工程策略明确了基于结构信息导向嵌入功能模块,以增加脱氨酶与非靶链结合力为核心思路。这不仅优化了底层的酶催化机制,也为后续可利用人工智能辅助的蛋白质结构预测和设计提供新的范式。随着AlphaFold等技术的快速成熟,未来多样化、个性化的碱基编辑器设计将更加高效精准,有望覆盖更多难治性遗传疾病的基因修复需求。
基于当前进展,ABE-NW及其衍生系列成为研发精准、高效、安全基因治疗新工具的代表。其简化且高效的开发流程也降低了实验阻力,加速了基因编辑器从实验室到临床的转化速度。展望未来,结构导向的模块化工程结合多样Cas9变体拓宽了靶点识别空间,提升了基因组编辑的应用范围。与此同时,通过不断优化编辑窗口和特异性,有望解决更多先天性疾病中的复杂单碱基变异,为个体化精准医疗奠定基础。 总结来说,这一简化的碱基编辑器工程策略以天然寡核苷酸结合模块为核心,开创了有效减少旁观编辑和脱靶风险的新途径。通过增强脱氨酶对DNA非靶链的结合特异性,成功缩小编辑窗口,实现了高效且精准的基因编辑,为解决遗传疾病的基因治疗难题提供了坚实的工具保障。
未来,结合先进的蛋白质工程和人工智能结构预测技术,这一策略将持续推动基因编辑技术的革新和应用,拓展精准遗传病治疗的可能性,成为基因技术发展史上的重要里程碑。 。









